North American Bean Thrips - Caliothrips fasciatus (Pergande) (Thysanoptera: Thripidae: Panchaetothripinae): A Perennial Export Problem for California Citrus Growers

Mark S. Hoddle, Ph.D.
Biological Control Specialist and Principal Investigator
Phone: (951) 827-4714
Fax: (951) 827-3086
mark.hoddle@ucr.edu

that are native to California.
Bean Thrips Biology and Reproduction
North American bean thrips (referred to here as bean thrips), Caliothrips fasciatus (Pergande) (Thysanoptera: Thripidae) are small insects (Figure 1) that were first collected from orange leaves in Yuba County California in 1894 (Bailey 1933). Bean thrips adults and larvae feed mainly on mature leaves and occasionally the skin of immature fruit. Female thrips deposit eggs into leaf material. Upon hatching from eggs, larvae feed and pass through two larval instars before reaching the propupal and pupal stages. As second instar larvae reach maturity they abandon host plants to fall into the soil where they transition into propupae and pupae. Following pupation, winged adults emerge from soil and likely fly back onto host plants to mate and feed. Sex determination for bean thrips is similar to other thrips, unfertilized eggs produce male thrips and fertilized eggs result in female offspring. Field populations of bean thrips have a sex ratio that is about 35% male and 65% female. Bailey’s (1933) breeding experiments in the lab indicated that female thrips that mate once and naturally live about 20 days stop producing female offspring after about 10 days and subsequent progeny are male only. If once-mated females naturally live for more than 50 days, female offspring production ceases after about 30 days and only males are produced thereafter. These results suggest that females need to mate frequently to continuously produce female offspring from fertilized eggs.
Bean Thrips Feeding Damage
Economic damage is caused by direct feeding injury by thrips larvae and adults. Bailey (1933) notes that larvae are responsible for more feeding damage than adults as they tend to be numerically superior on host plants, they are less active which results in larval feeding being limited to confined areas, and larvae tend to feed gregariously. Feeding damage is manifested as silvering on leaves, and when feeding damage is heavy, premature defoliation can occur. This has been observed for heavily infested pear trees and following defoliation events new growth and immature fruit are subjected to “sun-scald” over summer and the tree productivity is not as vigorous the following season (Bailey 1933). In the absence of leaves, thrips will feed on immature fruit which results in scarring and downgrading at packinghouses. Bailey (1933) notes that in once instance up to 15% of pear fruit were damaged at an orchard in the Berryesa Valley, Napa County California in 1930. Additional cosmetic damage results from the deposition of black fecal droplets on fruit surfaces by feeding larvae and adult thrips.
Bean Thrips Distribution Records
Bean thrips is native to California with a natural range that reportedly extends north into western Canada, south into Mexico, and east into Idaho and Florida (Mound et al. 2011; 2016). A record of this thrips from New York is based on one female (Wilson 1975). Part of the tentative non-native range of bean thrips may include several islands in Hawaii, (Hawaii, Lanai, Molokai, and Oahu), the Caribbean (Puerto Rico), and South America (Argentina, and Brazil) (Bailey1933; Mound et al. 2016). Examination of label data for the two slides in the US Museum of Natural History for bean thrips collected on Molokai indicated that the collections were made in 1943 from Argemone glauca, a native Hawaiian poppy. Similarly, specimens of C. fasciatus in the Hawaii Department of Agriculture collection were collected from A. glauca (collection data on one slide referencing “prickly poppy” could be the invasive weed Argemone mexicana), Sonchus oleraceus, and Chenopodium sp. are all from 1943-1946 (Janis Matsunaga, Hawaii Department of Agriculture, pers. comm. 3 Dec. 2019). Evidence of established breeding populations of bean thrips in Hawaii (Janis Matsunaga, Hawaii Department of Agriculture, pers. comm. 3 Dec. 2019) and the Caribbean are lacking (Dr. L.A. Mound, CSIRO Australia, pers. comm. 21 Oct. 2019 and Dr. C.A. O’Donnell, USDA-APHIS-PPQ, pers. comm. 28 Oct. 2019). Additionally, the few reports from Brazil and Argentina are likely misidentifications by non-specialists of C. phaseoli, a common pest thrips, for which there are no voucher specimens available for examination (Dr. A. Cavalleri, Universidade Federal do Rio Grande do Sul, Brazil, pers. comm. 22 Oct. 2019). Further, a 1927 record of C. fasciatus from China is likely erroneous and there is no conclusive evidence that bean thrips has established on citrus in Foochow (Fuzhou, Fujian Province) (Mound et al. 2011). Records of C. fasciatus from the Philippines, Spain, Italy, and “Palestine” are questionable (Bailey 1937). As Bailey (1933) notes, extensive surveys are required in the US (and from areas outside of North America for invasive populations) to determine the native range of C. fasciatus. For example, surveys have failed to find breeding populations of C. fasciatus in Australia (Hoddle et al. 2006). Putative species identifications resulting from surveys for C. fasciatus that are based on morphology can be readily confirmed with DNA analyses. In addition to C. fasciatus, two other species of Caliothrips are recorded from California, C. marginipennis (Hood) and C. phaseoli (Hood) (Mound et al. 2019). Globally, around 23 extant species of Caliothrips are recognized (ThripsWiki 2019).
The Quarantine Problem with Bean Thrips
So, why do we care as to where C. fasciatus is native or not native? In this instance, it’s about quarantine restrictions. Specifically, whether imports of California-grown navel oranges and mandarins are, for example, allowed into Australia and New Zealand, or they are denied entry and require fumigation if bean thrips are detected in shipments. The Australian Quarantine Inspection Service (AQIS) and Ministry of Primary Industries (MPI, New Zealand), have on numerous occasions found adult bean thrips overwintering in the “navels” of navel oranges and mandarins originating from California. The reason interceptions are of concern to AQIS and MPI is that bean thrips was once considered a serious agricultural pest in California attacking tree crops (e.g., avocados, pears, and walnuts), row crops (e.g., beans, peas, cantaloupes, and lettuce), cotton, and alfalfa. Economic damage consisted of premature defoliation due to heavy feeding on leaves, sunburn of branches exposed by defoliation events, scarring of fruit (15%-75% crop damage in pears), leaf silvering due to feeding, black spotting from fecal droplets, and plant death (notably of annuals) (Bailey 1937). Infestation severity varied from year to year and in very dry seasons damage could be widespread and severe on irrigated crops grown in interior regions (e.g., beans in Sutter County) (Bailey 1937). Hot weather and inadequate irrigation may have been, in part, significant factors driving severity of bean thrips infestations. Bailey (1938) notes that growing areas with summer temperatures averaging less than 18oC (64oF) rarely have problems with this pest.
Bean thrips commonly infests numerous weed species and native plants that grow in and around agricultural areas (Bailey 1933, 1937, 1938) (see Table 1 for putative plant list from which adults have been collected). The pest status of bean thrips has declined in California since the 1930’s. The reasons for this are unclear but could be due to better IPM practices and more efficacious pesticides, advances in irrigation technology, improved weed management, and development of resistant cultivars. Regardless of the current pest status of bean thrips in California, regulatory agencies, like AQIS and MPI, aim to prevent the establishment of pest species and inspections of imported produce are arguably the most important first step in managing the incursion of unwanted species.
Interestingly, overwintering bean thrips were first reported from navel oranges exported from California to Nebraska, Illinois, and Hawaii in 1899, 1907 and 1929, respectively (Bailey 1933; 1937). Bean thrips was not considered an economic pest of citrus at this time. However, mandatory fumigation of citrus fruit shipped to Hawaii was required between 1 September and 31 March each year (Bailey 1937). With respect to Australia, large volumes of navel oranges have been imported from California since the 1960’s and imports are arriving in Australia (and New Zealand) at a time of year (early to late summer in the southern hemisphere) that should be favorable for establishment as the climate would be hospitable and host plants in appropriate growth stages would be available. Yet despite extensive surveys in Australia, there is no evidence that bean thrips has established there (or New Zealand for that matter) (Hoddle et al. 2006).
Does the “Cold Stress Hypothesis” Explain Why Bean Thrips Has Failed to Establish in Areas Outside of its Native Range?
The question “why hasn’t bean thrips established in Australia or New Zealand?” despite repeated introductions into these countries may be answered, in part, by the “cold stress hypothesis.” Establishment of species outside of their native range is affected by many variables, of which important ones are climate (favorable in Australia [Hoddle et al. 2006] and probably in New Zealand too), food (abundant in both countries), and “propagule” pressure. Propagule pressure is a function of two things – frequency of introductions and numbers of individuals introduced each time during an incursion event. With respect to numbers of bean thrips detected during quarantine inspections by Biosecurity New Zealand personnel over the period 2014 – 2018, live bean thrips were found 35 times on citrus consignments imported from the USA. This included interceptions on oranges (25 live adults), lemons (5), mandarins (2), tangelos (2), and tangerines (1) (Alan Flynn, Biosecurity New Zealand, pers. comm. 25 November 2019).
It may be reasonable to assume that over the years there have been numerous introductions and overwintering thrips hiding inside “navels” have probably arrived undetected in Australia and New Zealand. Some of these thrips likely escaped detection during quarantine inspections and may have been delivered inside fruit to supermarkets, roadside stalls, and discarded within peels tossed onto compost heaps in backyard gardens. Assuming these thrips were alive at time of introduction, why did these escapees fail to establish in places with favorable climates and abundant food?
We are hypothesizing that bean thrips hasn’t established in areas where it is shipped because thrips surviving harvest, washing/waxing, cool storage, and refrigerated shipping are not “fit” when they arrive in a new locale. Thrips surviving transit inside navels may be weakened prior to harvest in California by accumulating winter cold stress. Following harvest, washing, waxing, and packing, thrips may experience more cold stress during post-harvest storage. For example, packed fruit may be held at 7oC [45oF] for several weeks if it’s not shipped promptly after picking. Once loaded onto a boat to Australia or New Zealand, thrips experience additional cold stress as fruit are stored at 3-4oC [37-39oF] for the 18-24 day transit period. Compounding cold stress resulting from winterization, storage, and shipping may have a significant negative impact on fitness, which is the longevity and fecundity, of surviving thrips. Simply put, cold-debilitated thrips that make it to Australia and New Zealand alive and escape into the wild are unable to found viable populations of sufficient size to overcome additional establishment barriers posed by natural enemies, competition from other thrips species, and stochastic environmental effects (e.g., a heat wave).
Another factor that needs consideration is possible reduction of female fertility that could result from cumulative cold stress. Female bean thrips that survive shipping to Australia and New Zealand may only be able to produce male progeny. As noted above, male progeny are produced from unfertilized eggs and female thrips need to mate regularly if they are to lay fertilized eggs that result in female offspring. If females that lay unfertilized eggs are unable to live long enough to mate with their sons they will be unable to lay fertilized eggs that result in daughters. High production of female progeny (i.e., daughters) is essential if incipient populations are to overcome establishment barriers.

the abdomen of a male bean thrips. Do these glands
release an aggregation pheromone?
Do Bean Thrips Use a Pheromone to Aggregate in Fruit?
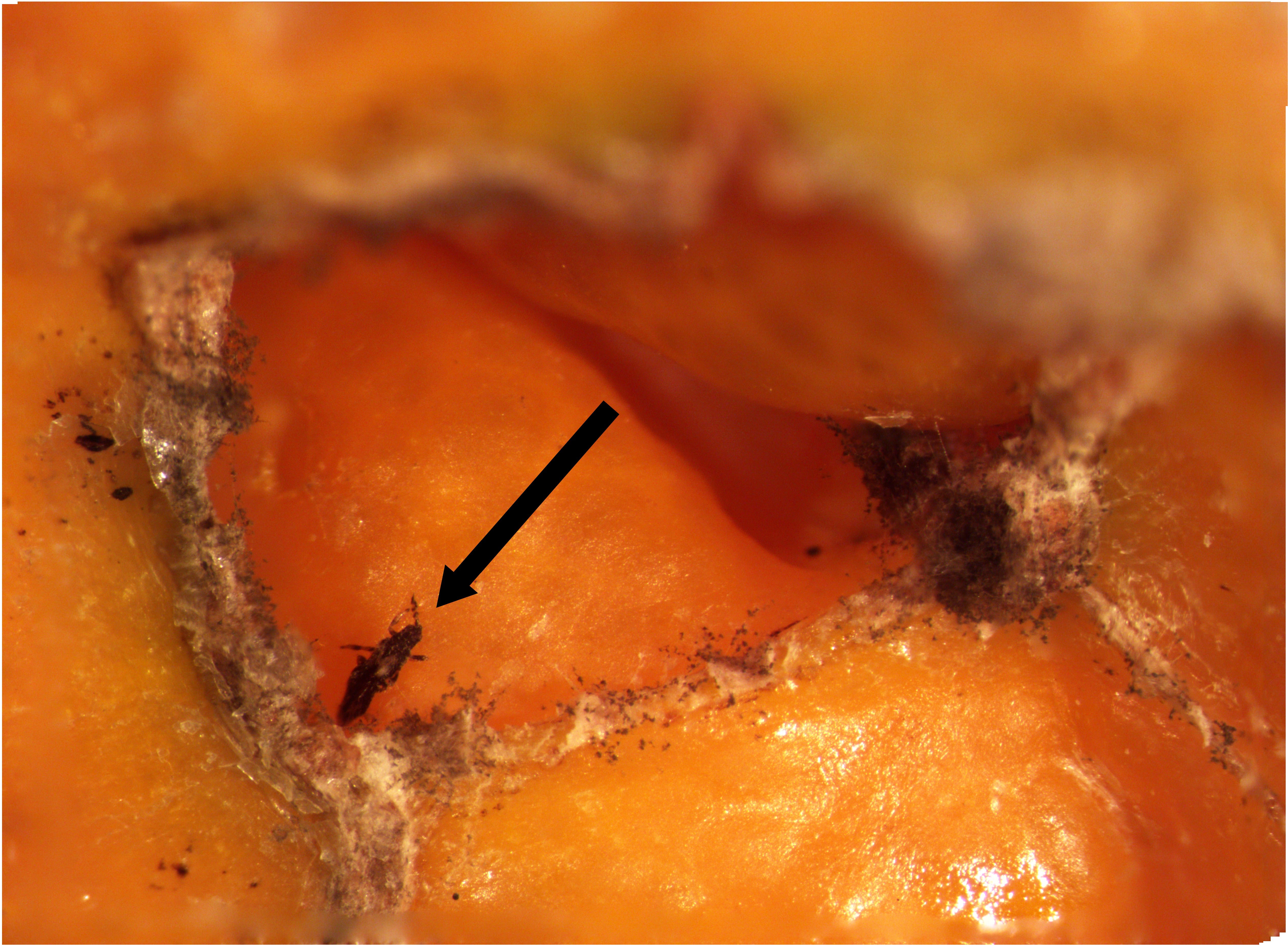
Unpublished data collected by Dr. Joseph Morse (Department of Entomology, UC Riverside) over winter 1997-1998 and 1999-2000 that measured levels of bean thrips infestations in navel oranges was passed to Mark Hoddle for analysis when Dr. Morse retired in 2018. The Morse lab cut open 7,300 navel oranges as they were harvested at survey orchards and around 3% had one or more live bean thrips inside the navel. The total number of bean thrips found in navels was around 500 and ranged from 1 to 40. Approximately 71% of infested fruit had one thrips, 10% had two, 4% had three, 3% had four, and the remaining 12% of infested fruit had five or more thrips sequestered inside navels. Preliminary statistical analyses looking for patterns of “clumping” indicated that bean thrips were not uniformly distributed in fruit and instead were highly aggregated. The negative binomial distribution can be used to assess clumping intensity, and a parameter, k, provides a measure of aggregation. Values of k around 2-3 suggest moderate clumping of samples. The tentative value of k from the Morse data set was > 30 suggesting extremely strong aggregation of bean thrips to relatively few navel oranges. Bailey (1933) also notes that bean thrips overwintering in field sites are often found hiding together in groups in protected spaces.
An important question to investigate is: “how do more than one bean thrips “decide” to collectively hibernate in a navel together when there are thousands of fruit hanging in trees that could be chosen?” We suspect that male bean thrips may release an aggregation pheromone from sausage-shaped glands located on the underside of the abdomen (Figure 2). The aggregation pheromone entices male and female thrips to enter the navel to communally overwinter. Increasing numbers of males in the navel could result in more pheromone wafting from an orange, and more thrips will be attracted to this individual fruit. Other species of thrips, like western flower thrips, Frankliniella occidentalis, a species native to California, produce pheromones, suggesting that pheromone use may be possible for bean thrips. Thrips pheromones operate over short distances (i.e., a few inches) and not over long distances (i.e., hundreds of yards) like sex pheromones produced by moths. Navel morphology may strongly influence selection of overwintering sites. Navels characterized by a small exterior opening that also penetrates deep into the fruit may be highly preferred by overwintering bean thrips (Morse pers. comm. 22 Oct. 2019). The possibility that bean thrips males produce an aggregation pheromone warrants investigation, because if it exists, a pheromone could provide new detection and management opportunities.
Where Do Bean Thrips That Infest Navels Come From?
Bean thrips adults have been recorded from a very large and diverse variety of plants (Table 1). Many potential feeding and breeding hosts are common weeds (e.g., prickly lettuce, Lactuca serriola) and native California plants that may be found in or around citrus orchards. Thrips are notoriously poor fliers and Bailey (1933) states that bean thrips are capable of only flying about 1-1.5 meters (3-4 feet) in a zig-zag or spiral pattern before landing. The pattern of dispersal in the field is irregular and is dependent on the direction and strength of the wind and the location of the source infestation (Bailey 1933). Because bean thrips are poor fliers incapable of controlled linear flight over long distances an effective cultural control practice to reduce the likelihood of adults reaching fruit to overwinter in may be good weed sanitation in orchards. Bean thrips infesting weeds on the orchard floor in close proximity to trees bearing fruit may simply need to fly a couple of feet to find overwintering sites (Figure 3). Weeding would remove potential host plants located within orchards, which are sometimes immediately under fruit bearing trees. Controlling weeds within 10-20 meters (30-60 feet or further from the boundary) of orchard margins could reduce migration rates into orchards and subsequent contamination of fruit in border rows. Orchards that have poor weed management tend to have high levels of bean thrips activity (Bailey 1933).

weeds growing on the orchard
floor into overhanging citrus canopies
to find overwintering sites?
Bean Thrips Surveys in Orchards Exporting Citrus
A two tier pre-clearance inspection system for navels grown in California and exported to Australia was implemented by USDA-APHIS in 2005. The first tier of the inspection begins with sticky card monitoring in navel orange orchards. Field experiments have documented that green colored stick cards are superior to yellow, white, or blue sticky cards for attracting bean thrips (Morse, 2006; Harmon et al. 2007). One trap is deployed every 5 acres of citrus and a log of captured bean thrips is updated weekly. If trap catches exceed 10 thrips per card then that block is ineligible for export to Australia. Prior to harvest, 50 randomly selected fruit are destructively sampled, and detection of bean thrips makes the orchard ineligible for export. The second tier of inspection occurs in the packing house where 175 oranges from grower lots are destructively sampled for bean thrips, and fruit are kept at 18.33oC for eight hours to encourage adult thrips movement from navels. Any contaminated lots are ineligible for export. As a consequence of increasing phytosanitary regulations, navel exports to Australia from California decreased by 65% over the period 1998-2005. Fruit that pass the pre-clearance are shipped within 3-4 days of harvest; the transit time by sea from California to Australia is 18-24 days, and fruit are stored at ~3oC in an unmodified atmosphere (B. Norby pers. comm. [Sunkist Growers] 2005). This monitoring and clearance program is no longer operating (Jim Craney, California Citrus Quality Council, pers. comm. 7 January 2020).
Conclusions
Bean thrips contamination of California-grown navel oranges and increasingly mandarins with navels that are exported is a perennial quarantine problem for citrus growers. There are no reliable reports of bean thrips having established populations outside of North America, the assumed native range. Consequently, there are no records of this thrips being a pest anywhere else in the world. In this regard, bean thrips appears to be a pest unique to California (and possibly a few other western states, like Idaho and Utah [Bailey 1937]) but only during the 1930’s. It is uncertain as to why bean thrips is no longer a significant agricultural pest in California (or elsewhere). One possible reason as to why bean thrips has failed to establish in countries receiving California citrus contaminated with bean thrips is because thrips have been debilitated by cumulative cold stresses that significantly reduces the fitness of survivors. The ability of bean thrips to form overwintering aggregations inside navels may be mediated by a male-produced aggregation pheromone. Elucidation of a male-produced aggregation pheromone could provide novel detection and control tools. The severity of bean thrips infestations of citrus fruit fluctuates from year to year, and annual variation may be driven, in part, by weed abundance which is affected by winter rainfalls and irrigation. Good weed abatement may reduce infestation risks to California citrus growers and subsequently lessen risks of shipment rejections at ports of entry.




Project Number
CRB Research Project #5500-210, “Mitigating Export Risks Associated with Bean Thrips”
Acknowledgments
Ruth Amrich, Lab Assistant IV, provided assistance with bean thrips colony maintenance and laboratory work, and summary data. Dr. Sandipa Gautam and Ping Gu generously provided bean thrips for some aspects of this work on the impacts of cold storage on bean thrips fitness traits. Ivan Milosavljević, Ph.D., Post-Doctoral Researcher, assisted with preliminary analyses of bean thrips aggregation data used in this article. Mike Lewis, Lab Assistant IV, provided photos. Dr. Joseph Morse provided bean thrips count data from cut fruit. Dr. Laurence A. Mound provided feedback on an initial draft of this article.
References
Bailey, S.F. 1933. The biology of the bean thrips. Hilgardia 7: 467–522.
Bailey, S.F. 1937. The Bean Thrips. Monograph Bulletin 609. University of California Experiment Station, Berkeley, USA.
Bailey, S.F. 1938. Thrips of economic importance in California. Circular of the University of California Berkeley Agricultural Experiment Station 346.
Harmon, J.A., Mao, C. X., Morse, J. G. 2007. Selection of colour sticky trap for monitoring adult bean thrips, Caliothrips fasciatus (Thysanoptera: Thripidae). Pest Management Science 63: 201-216.
Hoddle, M.S. 2020. North American bean thrips: A perennial export problem for California citrus growers. Citrograph 11(2): 40-43.
Hoddle, M.S.; Stosic, C.D.; Mound, L.A. 2006. Populations of North American bean thrips, Caliothrips fasciatus (Pergande) (Thysanoptera: Thripidae: Panchaetothripinae) not detected in Australia. Australian Journal of Entomology 45: 122-129.
Morse, J. G. 2006. Management of bean thrips. Citrus Research Board Annual Report. 3 pp. (last accessed 10 December 2019)
Mound, L.A.; Zhang, H.-R; Bei, Y.-W. 2011. Caliothrips tongi sp. n. (Thysanoptera: Thripidae) from China and a dubious record of North American bean thrips. Zootaxa 2736: 57-62.
Mound, L.A.; Nakahara, S.; Tsuda D.M. 2016. Thysanoptera-Terebrantia of the Hawaiian Islands: an identification manual. ZooKeys 549: 71–126.
Mound L.A.; Hoddle M.S.; Hastings A. 2019. Thysanoptera Californica - Thrips of California. Lucidcentral.org, Identic Pty Ltd, Queensland, Australia. Last accessed 22 October 2019.
ThripsWiki. 'Caliothrips
Wilson, T.H. 1975. A monograph of the subfamily Panchaetothripinae (Thysanoptera: Thripidae). Memoirs of the American Entomological Institute 23: 1–354.
Plants from which adult Caliothrips fasciatus have been collected
Table 1 below Listed plants are not necessarily breeding hosts for bean thrips. A true host plant would have bean thrips eggs, larvae, and adults living on it (larvae drop into the soil to pupate). This plant list was compiled from Bailey (1933; 1937; 1938), C. fasciatus collection data on slide mounted specimens in the Ewart thrips collection at UC Riverside, and field surveys in California by MSH. It is important to note that the taxonomy of listed plants may change and designations provided below were verified where possible using online resources.
|
Crop Plants |
|
|
|
Family |
Scientific name |
Common name |
|
Amaranthaceae |
Beta vulgaris L. |
Beets |
|
|
Beta vulgaris subsp. vulgaris |
Swiss chard |
|
|
Chenopodium album |
Pigweed |
|
|
Chenopodiastrum murale (L.) S. Fuentes, Uotila & Borsch |
Nettle-leaved goosefoot |
|
Amaryllidaceae |
Allium sativum L. |
Garlic |
|
Apiaceae |
Daucus carota sub species sativus (Hoffm.) Schübl. & G. Martens |
Carrot |
|
Asparagaceae |
Asparagus officinialis L. |
Asparagus |
|
Asteraceae |
Lactuca sativa L. |
Lettuce |
|
Brassicaceae (Cruciferae) |
Brassica oleracea L. |
Cabbage |
|
|
Brassica oleracea (L.) |
Cauliflower |
|
|
Brassica oleracea L. |
Kale |
|
|
Raphanus sativus L. |
Radishes |
|
|
Brassica rapa L. |
Turnips |
|
Cannabaceae |
Humulus lupulus L. |
Hops |
| Cannabaceae | Cannabis sativa subsp. indica (Afghan Indica Landrace) | Afghan Kush |
| Cannabaceae | Cannabis sativa subsp. sativa (African Malawi Sativa Landrace) | Malawi Gold |
|
Cucurbitaceae |
Citrullus lanatus (Thunb.) Matsum. & Nakai |
Watermelon |
|
|
Cucumis melo var. cantalupo Ser. |
Cantaloupe |
|
Cupressaceae |
Cupressus sp. |
Cyprus |
|
Ebenaceae |
Diospyros L. |
Persimmon |
|
Fabaceae |
Medicago sativa L. |
Alfalfa |
|
|
Phaseolus sp. |
Beans |
|
|
Trifolium pratense (L.) |
Clover, red |
|
|
Pisum sativum L. |
Peas |
|
Juglandaceae |
Juglans regia L. |
Walnuts |
|
Lauraceae |
Persea americana Miller |
Avocado |
|
Liliaceae |
Allium cepa L. |
Onions |
|
Malvaceae |
Gossypium hirsutum (L.) |
Cotton |
|
Moraceae |
Ficus carica L. |
Figs |
|
Musaceae |
Musa sp. |
Banana |
|
Oleaceae |
Olea europaea L. |
Olive |
|
Poaceae |
Zea mays L. |
Corn (young shoots) |
|
Rosaceae |
Prunus amygdalus Batsch |
Almond |
|
|
Malus sylvestris L. |
Apple |
|
|
Prunus persica Batsch |
Peach |
|
|
Pyrus communis L. |
Pear (especially Bartlett) |
|
|
Prunus domestica L. |
Prune |
|
Rutaceae |
Citrus latifolia Tanaka |
Lime |
|
|
Citrus sinensis (L.) Osbeck |
Orange |
|
|
Citrus reticulata (Blanco) |
Tangerine |
|
Solanaceae |
Capsicum sp. |
Bell pepper |
|
|
Solanum tuberosum L. |
Potato |
|
|
Lycopersicon esculentum P. Mill |
Tomatoes |
|
Vitaceae |
Vitis sp. |
Grape |
|
|
|
|
|
Ornamental and Wild Host Plants |
|
|
|
Adoxaceae |
Viburnum sp. |
Laurestine |
|
Amaranthaceae |
Amaranthus retroflexus L. |
Pigweed |
|
|
Salsoa kali L. |
Russian thistle or tumbleweed |
|
Asclepiadaceae |
Asclepias mexicana Cav. |
Milkweed |
|
Arecaceae |
Washingtonia filifera (Lindl.) H. Wendl. |
California fan palm |
|
Asteraceae |
Anthemis cotula L.
|
stinking chamomile or mayweed |
|
|
Aster sp. |
Asters |
|
|
Bidens pilosa L. |
Beggar's ticks or bur marigold |
|
|
Cirsium edule (Nutt.) |
Edible thistle |
|
|
Crepis sp. (L.) |
Hawksbeard |
|
|
Dahlia imperialis Roezl ex Ortgies |
Bell tree dahlia |
|
|
Echinacea purpurea (L.) Moench |
Eastern purple coneflower |
|
|
Erigeron canadensis (L.) Cronq. |
Horseweed |
|
|
Helianthus annuus L. |
Common sunflower |
|
|
Hemizonia sp. |
Tarweeds |
|
|
Lactuca serriola L. |
Prickly lettuce |
|
|
Lactuca serriola var. integrata Gren. & Godr. |
Prickly lettuce |
|
|
Pleurocoronis pluriseta (A.Gray) King & H.Rob. |
Bush arrowleaf |
|
|
Pseudognaphalium californicum (DC.) Anderb. |
California cudweed, California everlasting |
|
|
Sonchus oleraceus L. |
Sow thistle |
|
|
Stephanomeria sp. |
Wire lettuce |
|
Boraginaceae |
Heliotropium currassavicum L. |
Chinese pusley, salt heliotrope |
|
Brassicaceae |
Brassica rapa L. |
Common yellow Mustard |
|
Cannaceae |
Canna sp. (P. Mill) |
Wild canna |
|
Caryophyllaceae |
Stellaria media (L.) Vill. |
Chickweed |
|
Chenopodiaceae |
Atriplex sp. |
Saltbush |
|
|
Chenopodium murale L. |
Nettleleaf goosefoot |
|
Convolvulaceae |
Convolvulus arvensis L. |
Field bindweed |
|
Cucurbitaceae |
Echinocystis sp. |
Wild Cucumber |
|
Euphorbiaceae |
Ricinus communis L. |
Castor bean or castor oil plant |
|
Fabaceae |
Acacia sp. |
Wattles or acacias |
|
|
Acmispon americanus (Nutt.) Rydb |
Bird's-foot trefoil, deervetch, Spanish clover |
|
|
Acmispon glaber (Vogel) Brouillet |
Deerweed, California broom |
|
|
Caragana sp. |
Pea shrub |
|
|
Cassia sp. |
Golden cassia |
|
|
Indigofera decora (Lindl.) |
Spanish clover |
|
|
Leucaena leucocephala (Lam.) de Wit |
White leadtree |
|
|
Lupinus sp. |
Lupin |
|
|
Medicago polymorpha L. |
Bur clover |
|
|
Meliotus albus Medik. |
White sweet clover |
|
|
Pueraria montana (Lour.) Merr. |
Kudzu |
|
|
Wisteria sinensis |
Chinese wisteria |
|
|
Sophora sp. |
Kowhai |
|
|
Vicia sp. |
Wild vetch |
|
Fagaceae |
Quercus agrifolia Née |
California live oak |
|
|
Quercus lobata Née |
Valley oak |
|
Geraniaceae |
Erodium cicutarium (L.) L'Hérit. |
Stork's bill or filaree |
|
|
Geranium sp. |
Cranesbill |
|
Gramineae |
Arundinaria japonica (Sieb. & Zucc.) Mak. |
Bamboo |
|
|
Eleusine indica (L.) Gaertn. |
Wire grass |
|
Iridaceae |
Iris germanica L. |
Purple flag |
|
|
Gladiolus sp. |
Gladioli |
|
Lamiaceae |
Mentha sp. |
Mint |
|
Liliaceae |
Tulipa sp. |
Tulip |
|
Loasaceae |
Mentzelia laevicaulis (Dougl. ex Hook.) Torr. & Gray |
Blazing star |
|
Lythraceae |
Cuphea viscosissima (Jacq.) |
Tarweed |
|
Malvaceae |
Alcea rosea L. |
Hollyhock |
|
|
Malva parviflora L. |
Cheeseweed |
|
Melanthiaceae |
Chamaelirium luteum (L.) A. Gray. |
Blazing star |
|
Montiaceae |
Claytonia perfoliata Donn ex Willd. |
Miner’s lettuce |
|
Moraceae |
Ficus sp. |
Fig trees |
|
Musaceae |
Musa paradisiaca L. |
Banana |
|
Nyctaginaceae |
Mirabilis laevis (Benth.) Curran |
Wishbone bush |
|
Oleaceae |
Ligustrum sp. |
Privet |
|
Onagraceae |
Clarkia unguiculata Lindl. |
Elegant clarkia or mountain garland |
|
Papaveraceae |
Eschscholtzia californica Cham. |
California poppy |
|
Passifloraceae |
Passiflora mollissima (H.B.K.) Bail. |
Banana passionfruit, |
|
Plantaginaceae |
Digitalis sp. |
Foxglove |
|
Phrymaceae |
Mimulus sp. |
Monkey flower |
|
Pinaceae |
Pinus sp. |
Pine |
|
Poaceae |
Pseudosasa japonica (Siebold & Zucc. ex Steud.) Makino ex Nakai |
Arrow bamboo |
|
|
|
Unidentified grasses |
|
Polygonaceae |
Eriogonum fasciculatum Benth. |
California buckwheat |
|
|
Polygonum aviculare L. |
Knotweed, birdweed, pigweed |
|
Pteridaceae |
Adiantum sp. |
Maiden hair fern |
|
Rosaceae |
Coleogyne ramosissima Torr. |
Blackbrush |
|
|
Eriobotrya japonica (Thunb.) Lindl. |
Loquat |
|
|
Heteromeles arbutifolia (Lindl.) M. Roem. |
Toyon |
|
|
Rosa spp. |
Roses |
|
Ranunculaceae |
Anemone sp. |
Wind flowers |
|
|
Pyracantha sp. |
Firethorn |
|
Sapindaceae |
Aesculus californica (Spach) Nutt. |
California buckeye or California horse-chestnut |
|
Scrophulariaceae |
Verbascum virgatum Stokes ex with. |
Mullein |
|
Solanaceae |
Nicotiana glauca Graham. |
Tree tobacco |
|
Tropaeolaceae |
Tropaeolum majus L. |
Nasturtium |
|
Umbelliferae |
Foeniculum sp. |
Fennel |